Les abeilles sont impliquées dans le service écosystémique de pollinisation, aussi important pour la reproduction sexuée de la majorité des plantes à fleurs que pour de nombreuses productions agricoles
Chiffres clés sur la pollinisation animale
Près de 90 % des espèces de plantes à fleurs dans le monde sont pollinisées par des animaux (Ollerton et al. 2011), principalement des insectes tels que les abeilles, certaines espèces parmi les diptères (les syrphes par exemple), les coléoptères (les cétoines par exemple) ou les lépidoptères (les papillons de jour et de nuit). Dans les agrosystèmes, l’activité des pollinisateurs - qu’ils soient apportés artificiellement par l’homme en installant des colonies d’abeilles mellifères (Apis mellifera) par exemple, ou qu’ils proviennent naturellement du paysage environnant - contribue considérablement au rendement et à la qualité de nombreuses productions agricoles, principalement parmi les fruits, les légumes et les cultures oléo-protéagineuses. La production de semences de ces espèces mais aussi d’espèces fourragères (luzerne, trèfles) et de légumes dont on consomme les parties végétatives dépend également de la pollinisation animale. La synthèse de Klein et al. (2007) constitue l’étude la plus complète sur la dépendance de l’agriculture mondiale vis-à-vis de la pollinisation par les animaux (Fig. 1).

Cette synthèse montre que la production de fruits, de légumes et de graines de 87 cultures alimentaires dépend de - ou est améliorée par - la pollinisation animale (Fig. 2)

1 tandis que 28 cultures ne requièrent pas la pollinisation animale. Rapporté au tonnage, les cultures liées à la pollinisation animale représentent 35 % du volume de la production agricole dans le monde, sachant que les plantes les plus cultivées (telles que le blé, le maïs ou le riz) ne dépendent pas de la pollinisation animale (Klein et al. 2007). La valeur économique de la pollinisation animale a été estimée à 153 milliards d’euros pour l’année 2005 pour les cultures destinées à l’alimentation humaine (Gallai et al. 2009). Cette somme représente 9,5 % de la valeur de la production agricole mondiale pour ces cultures. C’est en illustrant le régime alimentaire de l’homme, avec et sans la présence des insectes, que l’on se rend compte de l’importance du service de pollinisation rendu (Fig. 3).

Pour la première fois en 2015, une grande revue de médecine a évalué la contribution de la pollinisation animale à la santé humaine et a mis en évidence que la baisse de diversité alimentaire associée au déclin des pollinisateurs pourrait causer des déficits en certains nutriments et engendrer une augmentation de la fréquence de certains troubles ou maladies (Smith et al. 2015).
Les abeilles, des super-pollinisateurs utilisés pour la pollinisation des cultures
Les abeilles sont considérées comme les principaux pollinisateurs au niveau mondial (Danforth 2006, Rader et al. 2015). Leur efficacité pollinisatrice est liée (1) à leur régime alimentaire exclusivement constitué de ressources tirées des fleurs (nectar et pollen, et parfois huiles florales), (2) aux poils branchus qui recouvrent leur corps et qui permettent une fixation et un transport efficaces des grains de pollen et (3) à la fidélité souvent observée d’un individu donné pour une espèce florale donnée au cours d’un même déplacement de butinage, voire au cours de sa vie. Pour assurer la présence d’abeilles dans les cultures à pollinisation entomophile (c’est-à-dire pollinisées par les insectes), deux approches peuvent être envisager : (1) l’apport direct d’abeilles par l’homme au sein des parcelles cultivées, (2) la gestion de l’environnement visant à promouvoir la présence des abeilles.
L’apport direct d’abeilles est la méthode la plus répandue pour satisfaire ce besoin en pollinisation entomophile dans les agrosystèmes (Aizen and Harder 2009). Distribuée mondialement depuis des décennies, l’abeille mellifère est aujourd’hui le principal pollinisateur de nombreuses cultures telles que celles d’amandes, de pommes ou de poires à travers le monde (Garibaldi et al. 2013). En plus d’avoir un effet positif sur la production agricole, cette pratique peut être bénéfique pour l’apiculture (production de miel et multiplication des colonies par exemple). D’autres espèces d’abeilles, à la fois des espèces sociales et des espèces solitaires, sont également utilisées (Table 1).

Les bourdons sont élevés et commercialisés à l’échelle mondiale depuis les années 1990

Apport de bourdons pour la pollinisation d’arbres fruitiers. La boîte contient trois colonies de l’espèce Bombus terrestris
(Fig. 4), notamment pour la pollinisation des tomates sous serre (Velthuis et van Doorn 2006). Le bourdon terrestre (Bombus terrestris) est en effet beaucoup plus efficace pour la pollinisation des tomates que l’abeille mellifère, car il pratique la « buzz pollination » ou pollinisation par bourdonnement (Banda et Paxton 1991). Ce succès a poussé les entreprises à promouvoir l’utilisation de B. terrestris pour la pollinisation d’autres cultures (pommes et poires par exemple) et également à investir dans l’élevage d’autres espèces de bourdons (Table 1).
Dans les régions tropicales et subtropicales, d’autres abeilles sociales sont utilisées pour la pollinisation des cultures. Les abeilles sans dard (« stingless bees » en anglais) sont considérés comme de très bons pollinisateurs des tomates sous serre (Melipona quadrifasciata par exemple), mais surtout de très bons pollinisateurs des cultures tropicales telles que l’avocat, la noix de coco, le café, la noix de macadamia ou la mangue. L’utilisation de quelques espèces d’abeilles solitaires pour la pollinisation s’est également développée depuis quelques dizaines d’années (Table 1), principalement l’osmie Osmia lignaria pour la pollinisation des vergers (Bosch et al. 2006) et la mégachile Megachile rotundata, massivement utilisée aux Etats-Unis et au Canada pour la pollinisation de la luzerne (Pitts-Singer & Cane 2011). On peut citer également différentes espèces d’Osmia pour la pollinisation des amandes, pommes, cerises et prunes, et Nomia melanderi pour la pollinisation de la luzerne (Pitts-Singer & Cane 2011).
Des utilisations à risque
La commercialisation d’un petit nombre d’espèces d’abeilles pour la pollinisation conduit à l’utilisation d’espèces hors de leur aire de distribution native. Ces pratiques impliquent des risques d’invasions biologiques, une des causes majeures de perte de biodiversité (Courchamp et al. 2017). C’est le cas par exemple de B. terrestris (Schmid-Hempel et al. 2014), utilisé mondialement pour la pollinisation des tomates sous serre, au sein de son aire de distribution naturelle (l’Europe) mais aussi au-delà, notamment en Amérique du Sud. Aujourd’hui, B. terrestris y est une espèce invasive causant le déclin des populations de bourdons natifs de cette région du globe. Les échanges commerciaux d’abeilles posent aussi la question de l’introduction de nouveaux parasites et pathogènes susceptibles d’affecter les populations d’abeilles sauvages.
L’intérêt des abeilles sauvages pour la pollinisation des cultures
L’apport d’abeilles élevées et commercialisées ne doit pas faire oublier le fort intérêt des populations sauvages d’abeilles présentes dans l’environnement. Plusieurs études mettent en évidence l’importance de la pollinisation assurée par les abeilles sauvages (par exemple Winfree et al. 2008, Tuell et al. 2009) ainsi que la complémentarité et la synergie d’action entre l’abeille mellifère et les abeilles sauvages, ainsi qu’entre les abeilles et les autres insectes pollinisateurs (par exemple Brittain et al. 2013, Garibaldi et al. 2013). Ceci s’explique par le fait que, selon leur morphologie et leur écologie, les pollinisateurs ont des comportements de visite des fleurs et/ou de récolte et de transport du pollen qui sont différents. La pollinisation par plusieurs espèces plutôt qu’une seule est de ce fait plus efficace. Par ailleurs, si l’on considère l’abeille mellifère, espèce la plus largement utilisée pour la pollinisation, (Aizen & Harder 2009) ont montré que la demande d’activité pollinisatrice pour l’agriculture augmente plus vite que ce que peuvent satisfaire les populations de cette espèce. Il apparaît donc essentiel, pour pérenniser le service de pollinisation et les rendements agricoles, d’assurer la présence à la fois de l’abeille mellifère et des abeilles sauvages, et plus globalement de l’ensemble de la faune pollinisatrice, dans les agrosystèmes (voir par exemple Isaacs & Kirk 2010, Chabert et al. 2015).
Le déclin des abeilles, une menace pesant sur le mariage « abeilles et agriculture »
La pollinisation par les abeilles constitue un service écosystémique fragile. Au cours des dernières décennies, plusieurs études ont mis en évidence un déclin des abeilles en Europe et en Amérique du Nord, constaté aussi bien sur la diversité des abeilles sauvages que sur les cheptels d’abeilles mellifères.
Déclin des abeilles sauvages
Comparativement à d’autres groupes d’insectes (les papillons ou les libellules par exemple), les abeilles sauvages sont mal connues, notamment en ce qui concerne leur distribution géographique, présente et passée. Cela est dû à leur grande diversité et à leur identification difficile, affaire de spécialistes (Requier & Le Féon 2017). Ces lacunes rendent difficile l’évaluation de l’état des populations. Par exemple, l’établissement de la récente liste rouge de l’UICN (Union Internationale pour la Conservation de la Nature) des abeilles d’Europe s’est heurté à ce manque de connaissances et pour plus de la moitié des espèces le risque d’extinction n’a pu être évalué (Nieto et al. 2014). Les données permettant de comparer l’état des populations à deux périodes distinctes sont donc rares mais quelques études existent toutefois pour l’Europe et les Etats-Unis. Par exemple, tirant parti de la forte tradition naturaliste en Grande-Bretagne et aux Pays-Bas, Biesmeijer et al. (2006) ont pu rassembler plusieurs dizaines de milliers d’observations d’abeilles sauvages dans ces deux pays, et mesurer l’évolution du nombre d’espèces d’abeilles présentes au sein de carrés de 10 km² entre deux périodes, avant et après 1980. Une diminution du nombre d’espèces a été observée dans 52 % des carrés étudiés en Grande-Bretagne et dans 67 % des carrés aux Pays-Bas. Parallèlement, le nombre d’espèces a augmenté dans seulement 10 % des carrés en Grande-Bretagne et 4 % des carrés aux Pays-Bas.
Déclin dans les cheptels d’abeilles mellifères
La production de miel et le nombre d’exploitations apicoles ont également subi une forte érosion dans de nombreux pays de l’Union Européenne et aux États-Unis en raison de l’augmentation des pertes de colonies. Dans ces pays, les pertes annuelles de colonies peuvent dépasser jusqu’à 60 % du cheptel d’un apiculteur, contre 5 % à 10 % avant l’apparition du syndrome d’effondrement (Potts et al. 2010a, Goulson et al. 2015). Ce syndrome d’effondrement comporte deux aspects : d’une part, une diminution de la survie des cheptels apicoles depuis les années 1990 allant jusqu’à doubler voire quadrupler le taux de mortalité hivernale selon les régions et les pays (vanEngelsdorp et al. 2008, Potts et al. 2010b, van der Zee et al. 2012, Chauzat et al. 2016, Seitz et al. 2016), d’autre part une soudaine disparition des abeilles adultes de leur ruche, apparu plus récemment en 2006 sans qu’aucune raison ne soit immédiatement perceptible, nommé CCD pour Colony Collapse Disorder (Oldroyd 2007).
Des causes communes aux déclins des abeilles sauvages et de l’abeille mellifère
Le maintien des abeilles dans un paysage requiert la présence de ressources florales et de sites et matériaux pour leurs nidifications (Requier & Le Féon 2017). Les abeilles peuvent donc être affectées par les facteurs de stress qui diminuent l’abondance et la diversité de ces deux types de ressources. Elles peuvent aussi être affectées plus directement par des pesticides ou des bio-agresseurs (qui comprennent les agents pathogènes, les parasites et les prédateurs).
Manque de ressources alimentaires ou de sites de nidification
Les changements d’occupation du sol et l’intensification de l’agriculture ont fortement modifié l’environnement conduisant à une simplification des paysages. L’augmentation de la taille des parcelles cultivées réduit les espaces interstitiels entre parcelles, sources de fleurs spontanées - aussi appelées adventices - et réduit aussi la quantité de sites de nidification (pour les abeilles terricoles, mais aussi cavicoles lorsque les haies sont supprimées par exemple). L’augmentation de l’usage d’intrants (fertilisants et pesticides) réduit la diversité floristique dans les prairies (sélectionnant les espèces les moins oligotrophes), induisant un effet négatif sur les abeilles, au premier rang desquelles les espèces oligolectiques qui ne peuvent se nourrir que sur un petit nombre d’espèces. Enfin, l’homogénéisation du type d’agriculture à l’échelle régionale (par exemple la céréaliculture avec les rotations de type colza-blé-tournesol) provoque une discontinuité dans les successions de floraison des cultures, conduisant les abeilles à subir des périodes où aucune culture n’est en fleur (Requier et al. 2015). Cette période de disette alimentaire s’avère être critique pour la survie des colonies d’abeilles mellifères en milieux agricoles, suite à un effet cascade de la disette alimentaire sur la dynamique démographique des colonies, les affaiblissant jusqu’à l’approche de l’hiver (Requier et al. 2016). Les changements climatiques peuvent également induire une réduction des ressources florales pour les pollinisateurs en causant une désynchronisation des dates de floraison des plantes et des périodes d’activité des insectes. Les abeilles les plus spécialisées (oligolectiques) seraient alors les plus menacées d’extinctions locales (généralement des abeilles dont la période de vol est courte et correspond à la période de floraison de leur plante(s) nourricière(s)).
Effets létaux et sublétaux des pesticides
Différentes familles de pesticides sont apparues successivement depuis la seconde moitié du 19ème siècle avec, dans plusieurs cas, des effets néfastes pour les colonies d’abeilles mellifères (voir par exemple un récapitulatif historique dans Albouy & Le Conte 2014). Depuis la fin des années 1990, les néonicotinoïdes tiennent le devant de la scène (Bruneau & Simon 2013). Ces insecticides sont neurotoxiques, c’est-à-dire qu’ils agissent sur le système nerveux des insectes. Ils subissent de nombreux tests avant d’être mis sur le marché, afin d’évaluer s’ils sont susceptibles de provoquer la mort de l’abeille mellifère, insecte non-cible (c’est-à-dire que l’on ne souhaite pas affecter par l’usage de ces insecticides) la plus utilisée dans les tests. Si l’insecticide provoque la mort de l’abeille dans les conditions d’utilisation prévues, il n’est pas mis sur le marché. Cependant, des effets sublétaux (c’est-à-dire ne provoquant pas la mort, mais modifiant la physiologie ou le comportement) peuvent exister (Desneux et al. 2007). En utilisant une technologie de pointe, le RFID (pour Radio Frequency IDentification), une étude a montré que les butineuses d’abeilles mellifères exposées au thiaméthoxame, molécule de la famille des néonicotinoïdes, voient leurs aptitudes à rentrer à la ruche diminuer (Henry et al. 2012). Ce trouble comportemental provoque donc la perte d’individus, ce qui affaiblit la colonie, la rendant plus sensible aux autres facteurs de stress (les bio-agresseurs par exemple). Qu’en est-il pour les abeilles sauvages ? Elles font l’objet de moins nombreuses études que l’abeille mellifère mais les effets néfastes n’en sont pas moins importants (Arena & Sgolastra 2014). Concernant plus spécifiquement les néonicotinoïdes, des études démontrent que leur utilisation affecte négativement la densité d’abeilles sauvages dans les paysages agricoles et diminue la croissance et la capacité reproductive des colonies de bourdons (Goulson et al. 2015, Rundlöf et al. 2015).
Bio-agresseurs
Dispersés géographiquement par les échanges commerciaux (notamment la commercialisation d’abeilles pour la pollinisation), les bio-agresseurs (agents pathogènes, parasites et prédateurs) affectent les abeilles partout dans le monde. La transmission d’agents pathogènes et de parasites se fait entre individus d’une même espèce le plus couramment (cas notamment des colonies d’abeilles mellifères regroupées au même endroit par les apiculteurs) ou entre individus d’espèces différentes, sur les fleurs par exemple. Ce phénomène est constaté entre l’abeille mellifère et les bourdons en particulier (Goulson et al. 2015).
Convergence vers une cause multifactorielle
Manque de ressources florales et de ressources pour la nidification, pesticides ou bio-agresseurs ? Si ces facteurs pris un par un peuvent avoir des effets négatifs sur les populations, ce sont les interactions entre ces facteurs de stress (Fig. 5)
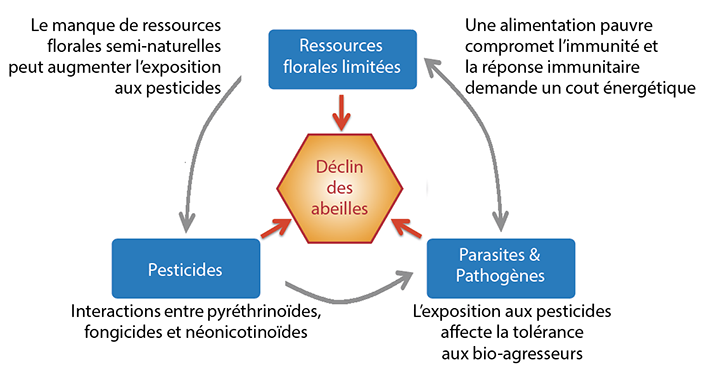
L’hypothèse la plus probante pour expliquer le déclin des abeilles porte sur des interactions multiples entre facteurs de stress comprenant les pesticides, le manque de ressources florales et les parasites et pathogènes.
qui constituent à l’heure actuelle l’explication la plus probante au déclin des abeilles (Potts et al. 2010a, Goulson et al. 2015). Par exemple, les effets négatifs d’un pesticide sont plus importants si l’abeille est conjointement affectée par un second facteur de stress, un autre pesticide ou un bio-agresseur par exemple (Goulson et al. 2015). La littérature scientifique fournit aujourd’hui de nombreux exemples d’interactions entre facteurs de stress : parasites × pathogènes (Retschnig et al. 2014), malnutrition × pathogènes (Di Pasquale et al. 2013), pesticides × pathogènes (Collison et al. 2015), pesticides × pesticides (Gill et al. 2012), et pesticides × climat ou contexte paysager (Henry et al. 2014).
Réconcilier « abeilles et agriculture » avec des actions de conservation
Le déclin des populations d’abeilles (l’abeille mellifère comme les espèces sauvages) observé en de nombreux points du globe depuis une trentaine d’années alerte les scientifiques et les pouvoirs publics par son ampleur écologique et socio-économique. La préservation des abeilles, de la filière apicole et du service de pollinisation est donc actuellement un enjeu majeur pour de nombreux acteurs (voir par exemple en France le plan national d’actions « France, terre de pollinisateurs » ou en Belgique le « Plan Fédéral Abeilles »). Dans les paysages agricoles, les facteurs de stress peuvent être nombreux et les bénéfices écologiques et socio-économiques de la présence des abeilles peuvent s’avérer considérables. C’est pourquoi des mesures en faveur des abeilles se développent, progressivement.
Comme exposé précédemment, le maintien des abeilles dans un paysage requiert la présence de deux types de ressources, des ressources florales et des ressources pour la nidification (Requier & Le Féon 2017). Parallèlement, les pesticides ou les bio-agresseurs peuvent affecter directement les populations. Les voies d’action en faveur des abeilles sont donc multiples. Il est nécessaire de mettre en place des mesures visant à maintenir ou augmenter les ressources florales et les ressources pour la nidification dans les paysages agricoles et à réduire l’usage des pesticides et la pression exercée par les bio-agresseurs.


Le ré-enrichissement des paysages agricoles en ressources florales est susceptible de prendre plusieurs formes. Les mesures les plus souvent mises en place actuellement consistent à restaurer et promouvoir les habitats semi-naturels riches en fleurs (Fig. 6) et les espaces semés avec des mélanges de plantes dites mellifères2 (Fig. 7) (Wratten et al. 2012). Augmenter la quantité d’habitats semi-naturels (haies, talus ou prairies par exemple) et œuvrer pour leur qualité (limiter les applications d’herbicides ou de fertilisants sur les bordures de champs par exemple) sont deux voies à privilégier. La flore sauvage présente dans ces habitats est plus susceptible de bénéficier à un grand nombre d’espèces d’abeilles, communes comme plus rares ou plus spécialisées dans leurs besoins alimentaires. Par ailleurs, ces habitats sont également plus enclins à fournir des sites de nidification (petites surfaces de sol nu, ronces ou branches mortes par exemple). Au contraire, les semis d’espèces mellifères (les jachères apicoles ou fleuries par exemple) n’apportent pas de ressources pour la nidification et sont susceptibles d’introduire dans l’environnement des plantes moins bénéfiques que les plantes sauvages, voire parfois néfastes dans le cas d’espèces exotiques et/ou invasives. D’autres mesures encouragent la ré-introduction de cultures entomophiles dans les systèmes culturaux, comme les légumineuses (telles que les luzernes, sainfoins ou trèfles) (Fig. 7), cultures fortement visitées par les abeilles (Rollin et al. 2013). La mise en place de ces cultures constitue une des mesures agri-environnementales les plus souvent choisies par les agriculteurs en France, du fait du faible coût des graines et de la facilité de leur intégration dans les systèmes de rotations culturales (Decourtye et al. 2011). Mais cette production fourragère n’a un réel intérêt durable que dans les régions où l’élevage est encore présent (Van Buskirk & Willi 2004). Enfin, la conservation des adventices constitue une autre mesure pertinente pour le maintien des abeilles dans les paysages agricoles (Fig. 7). Les adventices se révèlent être une ressource importante dans le régime alimentaire des abeilles (Requier et al. 2015). La conservation des adventices peu préjudiciables pour les cultures serait possible par une modification de certaines pratiques agricoles, en particulier une diminution de l’utilisation d’herbicides, telle que requise dans le cadre de la nouvelle législation européenne sur les pesticides (Barzman & Dachbrodt-Saaydeh 2011). Ces solutions prometteuses sont également bénéfiques à la conservation d’autres espèces impliquées dans les services écosystémiques tels que les prédateurs des ravageurs des cultures (Bianchi et al. 2006) et des espèces végétales rares indispensables à certaines espèces de pollinisateurs (Gibson et al. 2006).
Enfin, la diminution de l’usage des insecticides est bien évidemment recommandée pour préserver les abeilles. Qu’elle se fasse dans un contexte agronomique, ornemental ou « phobique », l’utilisation des insecticides doit être restreinte à des cas de nécessité extrême, et avec toutes les précautions suivantes : un contrôle des doses judicieux, une gestion des quantités utilisées également rigoureusement réfléchie et une pulvérisation une fois le soleil couché pour limiter l’exposition directe des abeilles et autres insectes floricoles diurnes (Decourtye et al. 2016).
Pour conclure
L’apport d’abeilles élevées et commercialisées est souvent envisagé pour assurer la pollinisation des cultures entomophiles. Cette approche ne doit pas faire oublier l’intérêt majeur des abeilles sauvages et des insectes pollinisateurs en général pour la stabilité et l’efficacité du service de pollinisation sur le long terme. Seule une réduction des facteurs de stress liés aux pratiques agricoles permettra de maintenir la diversité des pollinisateurs sauvages dans les paysages agricoles et conjointement d’assurer la bonne santé des colonies d’abeilles mellifères présentes.
Remerciements
Nous remercions chaleureusement les personnes ayant permis l’utilisation de leurs photographies : Marie Harruis, Nicolas Morison, Christophe Philippe et Bernard Vaissière. Nous remercions également Stan Chabert pour avoir partagé avec nous ses découvertes bibliographiques.
✻ INRA, UR 406 Abeilles et Environnement, Site Agroparc, 84914 Avignon, France
✻ Instituto de Investigaciones en Recursos Naturales, Agroecología y Desarrollo Rural (IRNAD), Sede Andina, Universidad Nacional de Río Negro (UNRN) y Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Mitre 630, CP 8400, San Carlos de Bariloche, Río Negro,
Argentina
Bibliographie
- Abrol D. (2010) Foraging behaviour of Apis florea F., an important pollinator of Allium cepa L.
- Journal of Apicultural Research, 49, 318-325.
- Arena M. & Sgolastra F. (2014) A meta-analysis comparing the sensitivity of bees to pesticides. Ecotoxicology, 23, 324-334.
- Albouy V. & Le Conte Y. (2014) Nos abeilles en péril. Editions Quae. 192 p.
- Aizen M.A.& Harder L.D.(2009) The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Current Biology, 19, 915-918.
- Banda H.J.& Paxton R.J.(1991) Pollination of greenhouse tomatoes by bees, Acta Horticulturae, 288, 194-198.
- Barzman M. & Dachbrodt-Saaydeh S. (2011) Comparative analysis of pesticide action plans in five European countries. Pest Management Science, 67, 1481-1485.
- Bianchi F., Booij C.J.H.& Tscharntke T. (2006) Sustainable pest regulation in agricultural landscapes : a review on landscape composition, biodiversity and natural pest control. Proceedings of the Royal Society B- Biological Sciences, 273, 1715-1727.
- Biesmeijer J.C., Roberts S.P.M., Reemer M., Ohlemüller R., Edwards M., Peeters T., Schaffers A.P., Potts S.G., Kleukers R., Thomas C.D., Settele J. & Kunin W.E.(2006) Parallel declines in
- pollinators and insect-pollinated plants in Britain and the Netherlands. Science, 313, 351-354.
- Bohart G.E.(1972) Management of wild bees for the pollination of crops. Annual Review of
- Entomology, 6029, 287-312.
- Bosch J., Kemp W.P.& Trostle G.E.(2006) Bee population returns and cherry yields in an orchard pollinated with Osmia lignaria (Hymenoptera : Megachilidae). Journal of Economic Entomology, 99, 408-413.
- Brittain C., Williams N., Kremen C. & Klein A.M. (2013) Synergistic effects of non-Apis bees and honey bees for pollination services. Proceedings of the Royal Society B, 280, 20122767.
- Bruneau E. & Simon N. (2013) Dépérissements et pesticides. Abeilles & Cie, 153, 27-32.
- Cane J.H.(2002) Pollinating bees (Hymenoptera : Apiformes) of U.S. alfalfa compared for rates of pod and seed set. Journal of Economic Entomology, 95(1), 22-27.
- Chabert S., Conjeaud J.-C. & Vaissière B. (2015). Production de semences hybrides. POLAPIS : vers une conduite raisonnée de la pollinisation entomophile. Bulletin Semences, 245, 47-51.
- Collison E., Hird H., Cresswell J. & Tyler C. (2016) Interactive effects of pesticide exposure and pathogen infection on bee health – a critical analysis. Biological Reviews, 91(4), 1006-1019.
- Courchamp F., Fournier A., Bellard C., Bertelsmeier C., Bonnaud E., Jeschke J.M. & Russell J.C.(2017) Invasion biology : specific problems and possible solutions. Trends in Ecology and Evolution, 31(1), 13-22.
- Danforth B.N., Sipes S.D., Fang J. & Brady S.G.(2006) The history of early bee diversification based on give genes plus morphology. Proceedings of the National Academy of Science (USA), 103(41), 15118-15123.
- Decourtye A., Mader E. & Desneux N. (2011) Landscape enhancement of floral resources for honey bees in agro-ecosystems. Apidologie, 41, 264-277.
- Decourtye A., Vidau C., Rollin O., Requier F., Rüger C., Allier F., Le Féon V., Kretzschmar A., Devilliers J., Henry M. & Odoux J.F.(2016) Fréquentation des cultures par les abeilles mellifères et sauvages : synthèse des connaissances pour réduire le risque d’intoxication aux pesticides. Cahiers Agricultures, 25(4), 44001.
- Desneux N., Decourtye A., Delpuech J.-M. (2007) The sublethal effects of pesticides on beneficial arthropods. Annual Review of Entomology, 52, 81-106.
- Di Pasquale G., Salignon M., Le Conte Y., Belzunces L.P., Decourtye A., Kretzschmar A., Suchail S., Brunet J.-L. & Alaux C. (2013) Influence of pollen nutrition on Honey Bee health : do pollen quality and diversity matter ? PLoS One, 8, e72016.
- Gallai N., Salles J.M., Settele J. & Vaissière B.E.(2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecological Economics, 68, 810-821.
- Garibaldi L.A., Carvalheiro L.G., Leonhardt S.D., Aizen M.A., Blaauw B.R., Isaacs R., Kuhlmann M., Kleijn D., Klein A.M., Kremen C., Morandin L., Scheper J. & Winfree R. (2014) From research to action : enhancing crop yield through wild pollinators. Frontiers in Ecology and the Environment, 12, 439-447.
- Garibaldi L., Steffan-Dewenter I., Winfree R., Aizen M.A., Bommarco R., Cunningham S.A, Kremen C., Carvalheiro L.G., Harder L.D., et al. (2013) Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science, 339, 1608-1611.
- Gibson R.H., Nelson I.L., Hopkins G.W., Hamlett B.J.& Memmott J. (2006) Pollinator webs, plant communities and the conservation of rare plants : arable weeds as a case study. Journal of Applied Ecology, 43, 246-257.
- Gill R.J., Ramos-Rodriguez O., Raine N.E.(2012) Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature, 491, 105–108.
- Goulson D., Nicholls E., Botias C. & Rotheray E.L.(2015) Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science, 347, 1255957.
- Heard T.A.(1999) The role of stingless bees in crop pollination. Annual Review of Entomology, 44, 183-206.
- Henry M., Beguin M., Requier F., Rollin O., Odoux J.F., Aupinel P., Aptel J., Tchamitchian S. & Decourtye A. (2012) A common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science, 336, 348-350.
- Henry M., Bertrand C., Le Féon V., Requier F., Odoux J.-F., Aupinel P., Bretagnolle V., Decourtye A. (2014) Pesticide risk assessment in free-ranging bees is weather and landscape dependent. Nature Communications, 5, 4359.
- Isaacs R. & Kirk A.K.(2010) Pollination services provided to small and large high bush blueberry fields by wild and managed bees. Journal of Applied Ecology, 47, 841-849.
- Klein A.M., Vaissière B.E., Cane J.H., Steffan-Dewenter I., Cunningham S.A., Kremen C. & Tscharntke T. (2007) Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society B, 274, 303-313.
- Klein A.M., Cunningham S.A., Bos M. & Steffan-Dewenter I. (2008) Advances in pollination ecology from tropical plantation crops. Ecology, 89, 935-943.
- Kuhn E.D.& Ambrose J.T.(1984) Pollination of ‘Delicious’ apple by megachilid bees of the genus Osmia (Hymenoptera : Megachilidae). Journal of the Kansas Entomological Society, 57, 169-180.
- Nieto A., Roberts S.P.M., Kemp J., Rasmont P., Kuhlmann M., Biesmeijer J.C., Bogusch P., Dathe H.H., De la Rúa P., De Meulemeester T., et al. (2014) European Red List of bees. Luxembourg : Publication Office of the European Union.
- Monzón V.H., Bosch J. & Retana J. (2004) Foraging behavior and pollination effectiveness of Osmia cornuta (Hymenoptera : Megachilidae) and Apis mellifera (Hymenoptera : Apidae) on ‘Comice’ pear. Apidologie, 35, 575-585.
- Oldroyd B.P.(2007) What’s killing American honey Bees ? Plos Biology, 5, 1195-1199.
- Ollerton J., Winfree R. & Tarrant S. (2011) How many flowering plants are pollinated by animals ? Oikos, 120, 321-326.
- Pitts-Singer T.L.& Cane J.H.(2011) The alfalfa leafcutting bee, Megachile rotundata : the world’s most intensively managed solitary bee. Annual Review of Entomology, 56, 221-237.
- Potts S.G., Biesmeijer J.C., Kremen C., Neumann P., Schweiger O. & Kunin, W. E.(2010a) Global pollinator declines : trends, impacts and drivers. Trends in Ecology & Evolution, 25, 345-53.
- Potts S.G., Roberts S.P.M., Dean R., Marris G., Brown M.A., Jones R., Neumann P. & Settele J. (2010b) Declines of managed honey bees and beekeepers in Europe. Journal of Apicultural Research, 49, 15-22.
- Rader R., Bartomeus I., Garibaldi L.A., Garratt M.P.D., Howlett B.G., Winfree R., Cunningham S.A., Mayfield M.M., Arthur A.D. et al. (2015). Non-bee insects are important contributors to global crop pollination. Proceedings of the National Academy of Sciences of the USA, 113, 146-151.
- Requier F., Odoux J.F., Tamic T., Moreau N., Henry M., Decourtye A. & Bretagnolle V. (2015) Honey bee diet in intensive farmland habitats reveals an unexpectedly high flower richness and a major role of weeds. Ecological Applications, 25, 881-890.
- Requier F., Odoux J.F., Henry M. & Bretagnolle V. (2016) The carry-over effects of pollen shortage decrease the survival of honeybee colonies in farmlands. Journal of Applied Ecology, doi : 10.1111/1365-2664.12836.
- Requier F. & Le Féon V. (2017) L’écologie des abeilles et ses enjeux pour l’agriculture. Abeilles & Cie, 176, 16-20.
- Retschnig G., Williams G.R., Mehmann M.M., Yañez O., de Miranda J.R.& Neumann P. (2014) Sex-specific differences in pathogen susceptibility in Honey Bees (Apis mellifera). PLoS One, 9, e85261.
- Rollin O., Bretagnolle V., Decourtye A., Aptel J., Michel N., Vaissière B.E.& Henry M. (2013) Differences of floral resource use between honey bees and wild bees in an intensive farming system. Agriculture, Ecosystems & Environment, 179, 78-86.
- Rundlöf M., Andersson G.K.S., Bommarco R., Fries I., Hederström V., Herbertsson L., Jonsson O., Klatt B.K., Pedersen T.R., Yourstone J. & Smith H.G.(2015) Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature, 521, 77-80.
- Schmid-Hempel R., Eckhardt M., Goulson D., Heinzmann D., Lange C., Plischuk S., Escudero L.R., Rahel Salath R., Scriven J.J.& Schmid-Hempel P. (2014) The invasion of southern South America by imported bumblebees and associated parasites. Journal of Animal Ecology, 83, 823-837.
- Sihag R.C.(1986) Insect pollination increases seed production in cruciferous and umbelliferous crops. Journal of Apicultural Research, 25, 121-126.
- Smith M.R., Singh G.M., Mozaffarian D. & Myers S.S.(2015) Effects of decreases of animal
- pollinators on human nutrition and global health : a modelling analysis. The Lancet, 386, 1964-1972.
- Tuell J.K., Fiedler A.K., Landis D. & Isaacs R. (2008) Visitation by wild and managed bees (Hymenoptera : Apoidea) to eastern US native plants for use in conservation programs.
- Environmental Entomology, 37, 707-718.
- Van Buskirk J. & Willi Y. (2004) Enhancement of farmland biodiversity within set-aside land. Conservation Biology, 18, 987-994.
- Velthuis H.H.W.& van Doorn A. (2006) A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie, 37, 421-451.
- Viana B.F., da Encarnacao Coutinho J.G., Garibaldi L.A., Braganca Gastagnino Guido L., Peres Gramacho K., da Silva F.O.(2014) Stingless bees further improve apple pollination and production. Journal of Pollination Ecology, 14, 261–269.
- Winfree R., Williams N.M., Gaines H., Ascher J. & Kremen C. (2008) Wild pollinators provide the majority of crop visitation across land use gradients in New Jersey and Pennsylvania. Journal of Applied Ecology, 45, 793-802.
- Wratten S.D., Gillespie M., Decourtye A., Mader E. & Desneux N. (2012) Pollinator habitat enhancement : benefits to other ecosystem services. Agriculture, Ecosystems & Environment, 159, 112-122.