La question des besoins nutritionnels de la colonie et des réserves disponibles est à la base de la compréhension du phénomène du cannibalisme du couvain. A la fois phénomène régulatoire et symptôme d’une modification de la dynamique des populations, le cannibalisme ne serait-il pas également dans certains cas un symptôme de dysfonctionnement ?
Circonstances du cannibalisme dans une colonie d’abeilles
En période de butinage empêché, les abeilles adultes peuvent pratiquer le cannibalisme sur certaines larves. Trois à cinq jours sans butinage, en période de pluie continue ou par grand vent, suffisent à déclencher le cannibalisme des larves de moins de 3 jours. Les ouvrières trouvent ainsi une source de protéines pour compenser la pénurie d’approvisionnement en pollen. Il s’agit en quelque sorte d’un système de régulation de l’offre et de la demande de pollen. Le couvain se conçoit alors comme une réserve de protéines. Dans cette pratique de cannibalisme, le flux trophallactique dans la colonie s’enrichit des protéines issues du tissu larvaire. Le niveau collectif de protéines est ainsi régulé, en réponse, en quelque sorte, à une consigne homéostasique circulant dans le superorganisme.
Comportement associé
En parallèle à ce comportement, les larves les plus âgées (de plus de 4 jours), dont l’élevage a constitué un investissement important en soins de la part des nourrices, sont quant à elles operculées plus tôt. Cette pratique n’est pas idéale pour le développement des larves mais permet de réduire la demande en pollen. La règle est simple : moins il y a de pollen stocké dans la ruche pendant le développement des larves, plus les larves sont operculées tôt. Bien que cela diminue la pression de la demande larvaire en situation d’urgence, ce mécanisme a un certain coût pour la colonie : les larves qui sont operculées avec un trop faible apport pollinique en subissent les conséquences après l’émergence, manifestant alors l’impact physiologique résultant d’une alimentation faible. Les poids corporels et les poids relatifs de ces abeilles sont bien inférieurs à la moyenne. Leur teneur en protéines est réduite, suggérant ainsi une durée de vie adulte plus courte.
Quelles conséquences pour la dynamique de la colonie ?
Ce comportement entraîne naturellement une perte de couvain et rapidement une diminution du nombre de nourrices. La dynamique peut s’avérer vite délétère si les conditions persistent à ne pas être favorables au butinage. La démographie larvaire change en effet en quelques jours. On sait que des larves moins nombreuses (cannibalisme), mal nourries (operculations précoces) et un couvain qui ne peut bénéficier de la température idéale (baisse de température par manque de nourrices) produisent vite des abeilles adultes avec des déficiences physiologiques. Les changements démographiques entraînent des perturbations dans la division du travail généralement bien équilibrée ainsi que dans l’allocation des ressources homéostasiques, ce qui peut à son tour influencer à nouveau la répartition démographique dans la colonie.
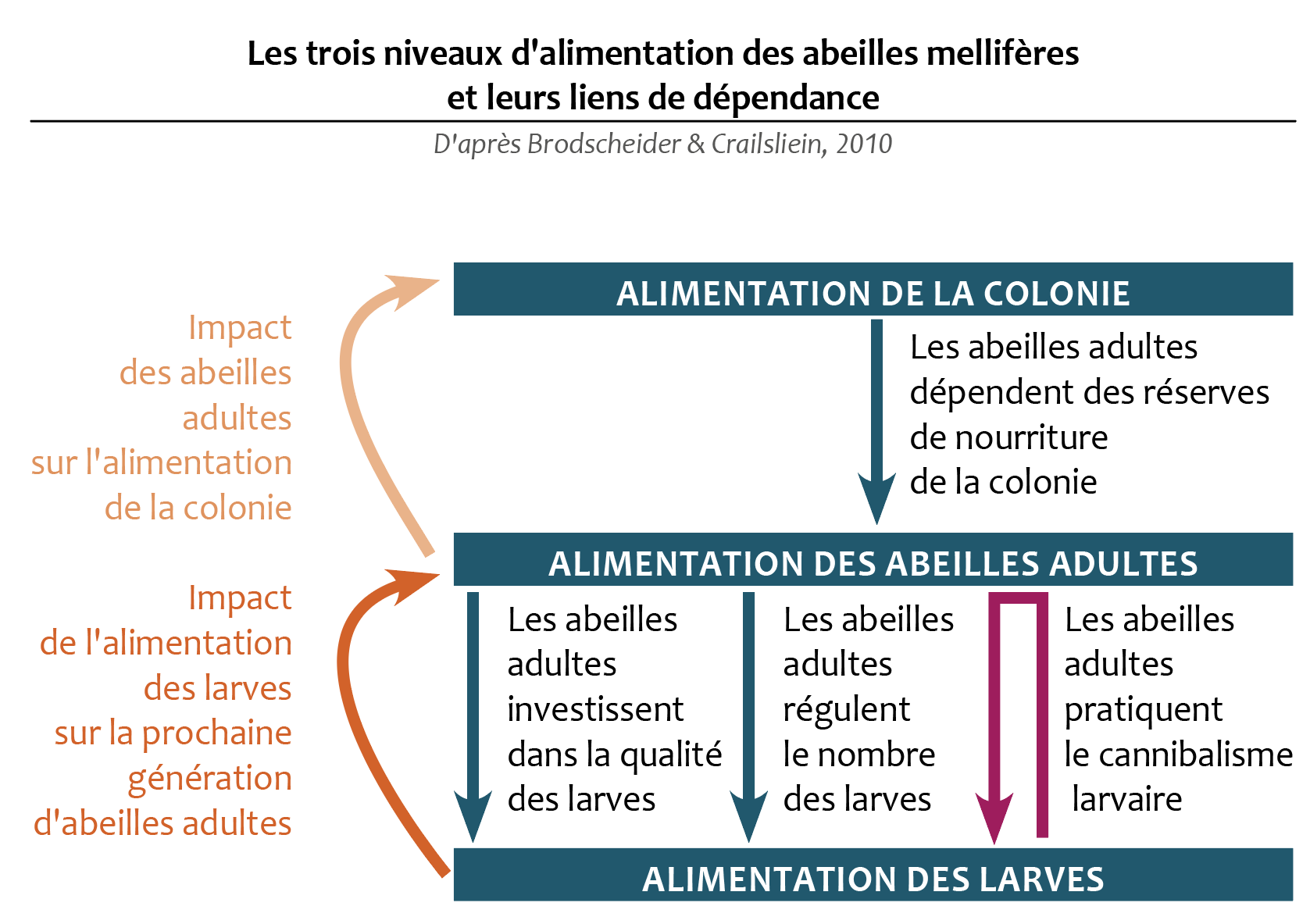
Selon Woyke, dans une étude déjà ancienne menée en 1976, des différences significatives ont été remarquées au niveau du taux de survie du couvain au printemps, en été et en automne en conséquence du cannibalisme. Le couvain a survécu jusqu’à l’émergence de l’adulte à 75 - 80 % au printemps, 80 - 90 % en été et 50 - 75 % en automne. Il s’agit naturellement d’estimations mais qui reflètent assez logiquement la question climatique qui sous-tend la pratique du cannibalisme de base. Le couvain de mâle a été davantage cannibalisé que le couvain d’ouvrière d’une manière générale, ce qui correspond à l’économie de la colonie. Au printemps et en été, les taux de survie du couvain de mâle et d’ouvrière restent similaires, que la colonie soit orpheline ou non. Par contre, en automne, un taux de couvain supérieur a survécu dans les colonies orphelines (75 %) par rapport aux colonies avec reine (50 - 65 %).
Concernant la ponte de la reine, les situations propices au cannibalisme ne la diminuent pas nécessairement significativement. Par contre, la reine réduit considérablement son déplacement. La ponte est donc localement concentrée, ce qui pourrait ainsi compenser les « trous » temporaires dans le couvain résultant du cannibalisme des jeunes larves.
Stock de nourriture et cannibalisme
Bien entendu, les abeilles font des réserves de nourriture et ces réserves doivent a priori permettre de réguler l’attribution des sources alimentaires dans la colonie même dans des conditions de butinage défavorables comme des périodes de pluie, de vent et de refroidissements. On sait bien sûr que le stock de pollen, principale réserve de protéines de la colonie, est accumulé à des niveaux bien plus bas que le stock de nectar. Il n’est disponible que sous la forme d’une réserve de quelques jours et il est vrai que la demande est particulièrement importante au moment où la colonie se développe de même que pour fabriquer des abeilles d’hiver. L’accumulation de pollen par temps favorable aurait un effet négatif sur le développement de la colonie. La colonie préférera donc constituer de faibles réserves de pollen pour gérer les urgences et mettre en place parallèlement des mécanismes compensatoires qui lui permettent une réaction rapide :
- Priorité à la recherche de pollen aux dépens de la recherche de nectar dès que possible (et frénésie de récolte pour arriver à une récolte de pollen conséquente en une seule journée) ;
- Réduction du niveau de couvain (et même arrêt de ponte dans certains cas) ;
- Réduction de l’alimentation du couvain ;
- Cannibalisme des œufs et des larves de 3 jours (avec une concentration probable sur les larves de mâles) ;
- Réaffectation des protéines pour alimenter les larves déjà bien nourries ;
- Maintien de l’alimentation de la reine à un niveau le plus constant possible.
Cannibalisme et santé de la colonie
Le cannibalisme larvaire est un mode de transmission horizontal des pathogènes. De ce fait, peut-il contribuer à une augmentation directe de la transmission des maladies du couvain dans la ruche ?
En 2021, la publication de Posada-Florez et al. a mis en lumière un fait important. Citons les auteurs : « Le virus de l’aile déformée (DWV), principal agent pathogène viral des abeilles mellifères associé à une mortalité accrue des abeilles mellifères et à la perte de colonies, est devenu très virulent avec la propagation de son vecteur, l’acarien ectoparasite Varroa destructor. La reproduction de varroa se produit dans les cellules de couvain operculé et les pupes infestées d’acariens dans ces cellules ont généralement des niveaux élevés de DWV. L’élimination des pupes infestées d’acariens par les abeilles ouvrières Varroa Sensitive Hygiene (VSH), conduit à la cannibalisation des pupes avec des charges DWV élevées, offrant ainsi une voie alternative de transmission du virus. »

La cannibalisation des pupes infectées par le virus des ailes déformées entraîne donc des niveaux élevés de ce virus chez les abeilles ouvrières. Le virus est ensuite transmis entre les abeilles de la colonie via la trophallaxie, permettant la circulation des variants du virus des ailes déformées qui se passe alors du vecteur acarien. Malgré les avantages connus du comportement hygiénique, il est possible que des niveaux très élevés d’activité VSH puissent entraîner une transmission accrue du DWV via le cannibalisme et la trophallaxie.
Les voies de transmission ont un impact sur la virulence et la génétique des agents pathogènes. Une connaissance approfondie de ces voies et leur contribution à la circulation des agents pathogènes est essentielle pour comprendre les interactions hôte-agent pathogène et concevoir des stratégies de contrôle. Cette publication récente, dont l’hypothèse reste à confirmer par d’autres études, devrait ouvrir la voie à d’autres recherches. Il serait intéressant d’étudier la question de l’impact du comportement de cannibalisme sur les autres maladies du couvain.
Le point de vue de l’apiculteur…
Témoignage de Philippe Aimé, apiculteur professionnel de Charente-Maritime
De mon point de vue, les tests de nettoyage destinés à juger du caractère hygiénique des colonies sont loin d’être anodins. Avec la mécanique de sélection mise en place en répandant la pratique du test hygiénique, on sélectionne des abeilles cannibales. On sélectionne les abeilles qui nettoient le plus vite. Cela ne fait que masquer l’expression de la maladie. Elles vont toutes cannibaliser, puisqu’elles nettoient en phase liquide. Le cannibalisme est un comportement limite, un mode de survie. On en fait ainsi peu à peu un mode de fonctionnement. La technique a son utilité, mais dans des cas particuliers, pas pour soutendre la construction de l’abeille.
Pour ma part, je cherche à ce que le nettoyage se fasse en phase solide, par extraction, pour que les ouvrières sortent le couvain malade. Il faut bien distinguer le cannibalisme du comportement de dépeçage. Le cannibalisme est la consommation du couvain. Les abeilles pratiquent une succion jusqu’à ce qu’il ne reste qu’une enveloppe sèche qu’elles ne peuvent pas retirer de la cellule. Le dépeçage c’est la suppression du couvain avec les mandibules.
Quel moyen les abeilles ont-elles de supprimer une prénymphe infectée ? Une prénymphe est à l’état liquide. Elles sont forcées de la cannibaliser. Quid de la répartition des charges virales ? Quel niveau de transfert de ces charges dans l’abeille ? Le cannibalisme conduit, via la trophallaxie, à transférer le virus des ailes déformées à toute la colonie par exemple. Une étude l’a mis à jour en 2021. Il ne faut pas exacerber le cannibalisme par la sélection issue du test hygiénique.
Comment repérer des abeilles cannibales ? Je prends une prénymphe ou une nymphe que je pose sur la tête d’un cadre. Je regarde ensuite à quelle vitesse les abeilles se précipitent dessus. J’observe aussi comment elles le font. Avec la langue ? Avec les mandibules pour attraper et tirer ? Je reste fasciné par le degré d’incompréhension de ce qu’il se passe dans la ruche malgré toutes les techniques d’observation dont nous disposons aujourd’hui. Il est devenu impératif de retrouver une compréhension du fonctionnement de la colonie d’abeilles.
Dans un même rucher, on peut observer des colonies avec des comportements très différents concernant les rentrées de pollen. Certaines colonies déviantes ne rentrent pas de pollen ou surconsomment et ont un besoin en protéines supérieur à leur capacité de collecte. Elles pratiquent un cannibalisme déviant. Le comportement normal d’une colonie en période de disette, c’est d’abord le retrait, le cannibalisme du couvain de mâles. Puis éventuellement un arrêt de ponte. Aujourd’hui, du fait d’une sélection contestable, on se trouve avec des abeilles productrices d’abeilles et plus des abeilles productrices de miel. Ce sont des colonies qui font du couvain en permanence. La conséquence c’est un beau gâchis : la reine pond et les ouvrières éliminent le surplus de leur côté. On se retrouve face à une double destruction : la reine vide sa spermathèque d’un côté et le couvain est supprimé de l’autre pour réguler la population.
Le cannibalisme est un phénomène instable, peu ou pas lié à la gestion de varroa. Je prends l’exemple d’une de mes colonies. A un an d’intervalle (de septembre à septembre), sans changement de reine, cette colonie est passée d’un cannibalisme exacerbé avec une charge varroa élevée (7 varroas pour 100 abeilles) à, 12 mois plus tard, le statut de colonie nettoyeuse par extraction mandibulaire avec une charge en varroas de l’ordre de 0,2/0,3 varroas pour 100 abeilles. Je pensais que cette colonie serait morte à la fin du premier hiver. Elle est morte en fin du deuxième hiver et a toujours été sous assistance alimentaire hivernale.
Il semblerait, d’après mes observations, que le cannibalisme soit effectué par des abeilles jeunes (avant la fonction cirière). Cela génère une désorganisation castique et des colonies très consommatrices de cire.
Je crois sincèrement que le monde apicole empêche les abeilles de s’en sortir. Le cannibalisme est au cœur du problème de l’absence de résilience de l’abeille. Il faut réorganiser les pratiques et retourner au temps long des abeilles. Ce n’est pas parce qu’on voit la maladie que la colonie ne va pas s’en sortir. Inversement, ce n’est pas parce qu’on ne voit pas les symptômes que la colonie n’est pas malade.
Avec mon expérience empirique, j’ai tendance à dire : ne rien faire à l’abeille c’est faire beaucoup pour elle.
Références :
Blaschon, B., Guttenberger, H., Hrassnigg, N., & Crailsheim, K. (1999). Impact of bad weather on the development of the broodnest and pollen stores in a honeybee colony (Hymenoptera : Apidae). Entomologia Generalis, 49-60.
Defèr, A. Classification of Honeybee Larval Stages Using CNNs Applied to Image Data.
O’Shea-Wheller, T. A., Rinkevich, F. D., Danka, R. G., Simone-Finstrom, M., Tokarz, P. G., & Healy, K. B.(2022). A derived honey bee stock confers resistance to Varroa destructor and associated viral transmission. Scientific Reports, 12(1), 1-19.
Posada-Florez, F., Lamas, Z. S., Hawthorne, D. J., Chen, Y., Evans, J. D., & Ryabov, E. V.(2021). Pupal cannibalism by worker honey bees contributes to the spread of deformed wing virus. Scientific Reports, 11(1), 1-12.
Robinson, G. E., Page, R. E., & Fondrk, M. K. (1990). Intracolonial behavioral variation in worker oviposition, oophagy, and larval care in queenless honey bee colonies. Behavioral ecology and sociobiology, 26(5), 315-323.
Rodriguez‐Rodriguez, M., & Kang, Y. (2016). Colony and evolutionary dynamics of a two‐stage model with brood cannibalism and division of labor in social insects. Natural Resource Modeling, 29(4), 633-662.
Rowland, B. W., Rushton, S. P., Shirley, M. D., Brown, M. A., & Budge, G. E.(2021). Identifying the climatic drivers of honey bee disease in England and Wales. Scientific reports, 11(1), 1-10.
Schmickl, T., & Crailsheim, K. (2007). HoPoMo : A model of honeybee intracolonial population dynamics and resource management. Ecological modelling, 204(1-2), 219-245.
Schmickl, T., & Karsai, I. (2017). Resilience of honeybee colonies via common stomach : A model of self-regulation of foraging. PloS one, 12(11), e0188004.
Schmickl, T., & Crailsheim, K. (2004). Inner nest homeostasis in a changing environment with special emphasis on honey bee brood nursing and pollen supply. Apidologie, 35(3), 249-263.
Schmickl, T., & Crailsheim, K. (2001). Cannibalism and early capping : strategy of honeybee colonies in times of experimental pollen shortages. Journal of Comparative Physiology A, 187(7), 541-547.
Schmickl, T., & Crailsheim, K. (2002). How honeybees (Apis mellifera L.) change their broodcare behaviour in response to non-foraging conditions and poor pollen conditions. Behavioral Ecology and Sociobiology, 51(5), 415-425.
Schmickl, T., Blaschon, B., Gurmann, B., & Crailsheim, K. (2003). Collective and individual nursing investment in the queen and in young and old honeybee larvae during foraging and non-foraging periods. Insectes Sociaux, 50(2), 174-184.
Soroker, V., Erez, T., Bonda, E., Cahanov, P., Rueppell, O., Wagoner, K., & Chejanovsky, N. The Interactions between Hygienic Behavior, Immune Response, and Virus Infection in Honeybees. Immune Response, and Virus Infection in Honeybees.
Spivak, M., & Danka, R. G.(2021). Perspectives on hygienic behavior in Apis mellifera and other social insects. Apidologie, 52(1), 1-16.
Thaduri, S. (2019). Virus dynamics in naturally Varroa-resistant honeybee populations. pub.epsilon.slu.se
Traynor, K. S., & Lamas, Z. S.(2021). Social disruption : Sublethal pesticides in pollen lead to Apis mellifera queen events and brood loss. Ecotoxicology and Environmental Safety, 214, 112105.
Woyke, J. (1977). Cannibalism and brood-rearing efficiency in the honeybee. Journal of Apicultural Research, 16(2), 84-94.