Le développement saisonnier
Le cycle biologique de la colonie est réglé par la ponte de la reine qui démarre plus ou moins précocement à la fin de l’hiver et au début du printemps en fonction de plusieurs para mètres parmi lesquels se trouvent les écotypes d’abeilles et le climat. Le pic de population d’abeilles dans la colonie est atteint en mai-juin et leur nombre baisse progressivement à partir du mois de juillet. En fin de saison, la ponte dépend fortement des températures de l’arrière-saison avant d’arriver à l’hivernage où une grappe d’abeilles d’hiver protège la reine. Ces abeilles d’hiver sont bio logiquement différentes des abeilles d’été avec des corps gras plus développés, un taux plus élevé de protéines dans l’hémolymphe et dans les glandes hypopharyngiennes, un taux inférieur d’hormone juvénile. Ces différences sont des traits de longévité. La colonie constitue sa réserve d’abeilles d’hiver dès le déclin de la ponte de la reine. Il s’agit d’une sorte de régulation hormonale interne. Moins de couvain permet par exemple aux nourrices de stocker la vitellogénine (protéine de longévité base de la gelée royale) dans leurs corps gras.
Le climat et le développement de la colonie
Naturellement, les variations climatiques saisonnières ont un impact sur le développement de la colonie. Des vagues de froid au printemps au moment du développement de la ponte de reine ou des arrière-sai sons plus chaudes qui suscitent une poursuite de la ponte ont des consé quences sur le développement géné ral de la colonie. Les pics climatiques, caractérisés par des épisodes brutaux et extrêmes (gels, pluie, sécheresse), ne sont pas sans conséquences sur la vie de la colonie dans le sens où ils perturbent son développement, influençant les entrées de pollen et de nectar, modifiant l’investissement de l’énergie dépensée et facilitant le développement de certains pathogènes. La dynamique de la population d’abeilles dans la colonie est soumise plus ou moins directement à ces aléas climatiques.
La dynamique de la population
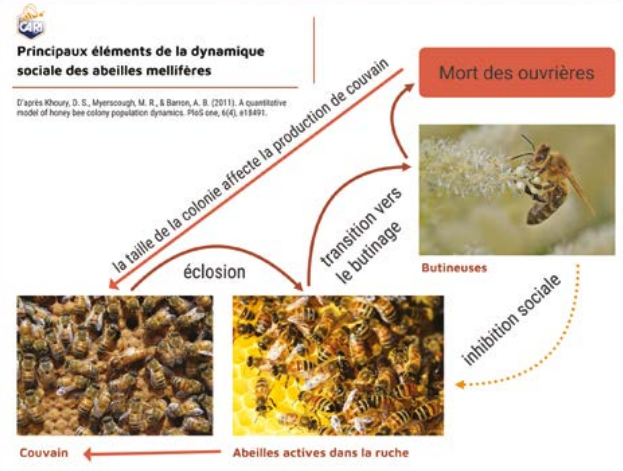
Une colonie d‘abeilles mellifères est une population d‘individus apparentés qui interagissent. On se trouve face à une société très complexe influencée par une dynamique complexe de la population. Les individus du superorganisme ont des rôles ou des affectations qui évoluent au sein du groupe tout au long de leur vie. Leur durée de vie est fortement influencée par leur rôle. On sait que la division du travail entre la population d’ouvrières dépend de l‘âge et des besoins de la colonie. Selon un principe général, les jeunes ouvrières sont plutôt employées dans la ruche à des tâches d‘entretien de la colonie et de soins au couvain comme le nourrissage. Ce ne sont que les ouvrières plus âgées qui se chargent de l’approvisionnement en nourriture de la colonie et qui ont un contact avec l’environnement extérieur via le butinage. Ce principe répond à un processus de développement basé sur un comportement social utilitaire. Ceci explique que ce grand principe peut être remis en question selon les besoins de la colonie. Si les butineuses voient leur nombre baisser drastiquement sous l’effet d’un stress environnemental par exemple, les autres ouvrières de la colonie vont accélérer leur développement comportemental pour entrer dans une dynamique de compensation : elles vont donc butiner précocement et sans doute mourir précocement. A l’inverse, s’il y a une surabondance de butineuses et un manque de nourrices, le développement comporte mental de certaines butineuses peut régresser et elles peuvent réendosser le rôle de nourrices. On parle d’inhibition sociale. L’ensemble de ces adaptations comportementales est régi par un mécanisme phéromonal désormais bien identifié.
Une interaction claire existe entre l’affectation des ouvrières de la colo nie et leur longévité. Si les ouvrières commencent à butiner tôt pour pal lier au manque de butineuses, leur durée de vie risque d’en être réduite et le temps consacré au couvain est également réduit, ce qui peut avoir un impact important sur la croissance de la colonie. De même, les différents stress bien connus (maladies, varroa, etc.) peuvent impacter la croissance des colonies et conduire à l’affaiblissement des populations d’ouvrières.
Bibliographie
Winston ML (1987) The biology of the honey bee. Cambridge : Harvard University Press.
Seeley TD (1982) Adaptive significance of the age polyethism schedule in honeybee colonies. Behavioral Ecology and Sociobiology 11 : 287–293.
Huang Z-Y, Robinson GE (1996) Regulation of honey bee division of labor by colony age demogra phy. Behavioral Ecology and Sociobiology 39 : 147–158.
Robinson GE, Page RE, Strambi C, Strambi A (1992) Colony integration in honey bees : mechanisms of behavioural reversion. Ethology 90 : 336–350.
Le Conte, Y., & Navajas, M. (2008). Climate change : impact on honey bee populations and diseases. Revue Scientifique et Technique-Office International des Epizooties, 27(2), 499-510.
Leoncini I, Le Conte Y, Costagliola G, Plettner E, Toth AL, et al. (2004) Regulation of behavioral maturation by a primer pheromone produced by adult worker honey bees. Proceedings of the National Academy of Sciences of the United States of America 101 : 17559–17564.
Rueppell O, Kaftanouglu O, Page RE (2009) Honey bee (Apis mellifera) workers live longer in small than in large colonies. Experimental Gerontology 44 : 447–452.
Khoury, D. S., Myerscough, M. R., & Barron, A. B.(2011). A quantitative model of honey bee colony population dynamics. PloS one, 6(4), e18491.